Forschung
pathogene Pilze und Oomyzeten
Botrytis cinerea ist ein nekrotropher Pilz, der eine Vielzahl von Pflanzen befallen kann, darunter viele wichtige Kulturpflanzen. Neben phytotoxischen Proteinen, die zur Ausprägung der Infektion beitragen, setzt dieser Pathogen auch kleine RNAs gegen die Pflanze ein. Das Phänomen des Cross-Kingdom RNAi als natürliche Infektionsstrategie wurde erstmals in diesem Pathogen nachgewiesen (Weiberg et al. 2013).

Botrytis Grauschimmel auf verschiedenen Früchten.
Hyaloperonospora arabidopsidis ist ein obligat-biotropher Parasit, der zur Klasse des falschen Mehltaus (Peronosporales, Oomycetes) gehört. Dieser Pathogen hat sich hochgradig auf die Besiedlung von Arabidopsis thaliana spezialisiert. Dafür bildet Hyaloperonospora Infektionsstrukturen aus, sog. Haustorien, worüber Effektormoleküle in die Pflanzenzellen eingeschleust werden. Auch dieser Pathogen benutzt Cross-Kingdom RNAi als Infektionsstrategie (Dunker et al. 2020).
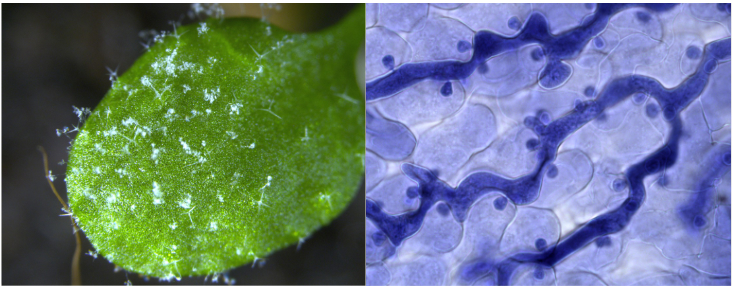
Links: Arabidopsis falscher Mehltau, rechts: Hyaloperonospora Haustorien während der Besiedlung eines Arabidopsis Keimlings.
Cross-kingdom RNAi
Cross-kingdom RNAi
Mikrobielle Pathogene schleusen kleine regulatorische RNAs in ihre Wirtspflanzen ein, um die Immunantwort zu unterdrücken. Dabei binden die pathogenen RNAs an Argonaute (AGO) Proteine der Pflanze und bilden sogenannte RNA-Induced Silencing Komplexe (RISCs), die sequenz-spezifisch die Translation von pflanzlichen mRNAs unterbinden. Diesen Virulenz-Mechanismus nennt man Cross-kingdom RNAi (Weiberg et al. 2014, Dunker et al. 2020).
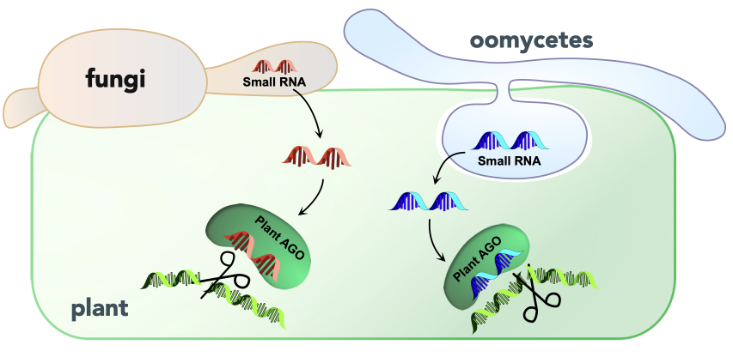
Cross-kingdom RNAi Reporter
Wir entwickeln neuartige Technologien, um den Transfer von RNAs zwischen Wirt und Pathogenen, ihre Wirkungsweise und ihr Ausmaß besser zu verstehen. Dazu haben wir Cross-kingdom RNAi Reporter-Systeme etabliert, die uns über Markergen-Expression die Übertragung von pathogenen RNAs in ihren Wirtspflanzen in zeitlicher und räumlicher Auflösung anzeigen. Diese Information können wir z.B. über Fluoreszenz-Imaging in live erzeugen und so verstehen, in welchem Infektionsstadium pathogene RNAs übertragen werden (Cheng et al., 2023).
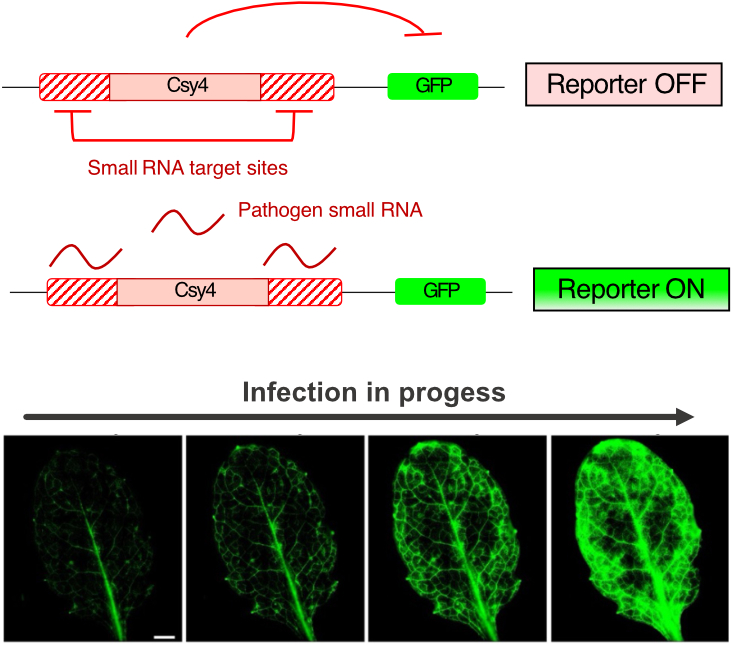
Oben: Schema eines cross-kingdom RNAi reporter contructs für die Expression in Pflanzen. Csy4 codiert eine RNA Endonuklase, die über eine spezifische RNA Erkennungssequenz (Csy4-binding site) an ein GFP Reportergen gekoppelt ist. Die Csy4 Kassette ist mit Target-Sequenzen von ausgewählten Pathogen RNAs fusioniert. Wenn der Pathogen diese kleinen RNAs während der Infektion in die Reporterpflanze einschleust, wird die Expression des Csy4 Gens abgeschaltet und GFP Expression aktiviert (siehe unten).
Extrazelluläre Vesikel
Wie werden RNAs von Pathogenen in ihre Wirtspflanzen übertragen? Extrazelluläre Vesikel (EVs) sind 50-500 nm große Lipid-Nanopartikel, die von mikrobiellen Pathogenen und Pflanzen abgesondert werden. EVs können mit verschiedenen Proteinen und RNAs beladen sein (Ruf et al. 2022). Der Pilz Botrytis cinerea benutzt EVs zur Übertragung von kleinen RNAs in die Wirtspflanze. Hierbei erforschen wir die molekularen und zellulären Zusammenhänge der EV-Biosynthesis, der RNA-Beladung und der Aufnahme von EVs und extrazellulärer RNA durch Organismen.
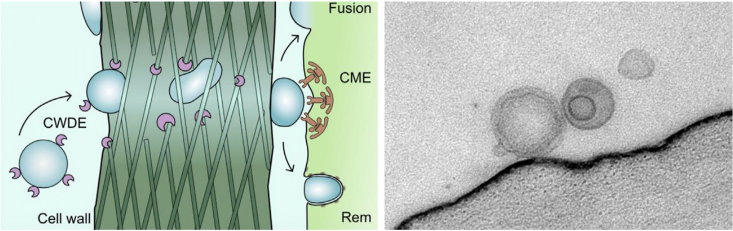
Links: Pathogen EVs müssen die pflanzliche Zellwand überwinden, um RNAs in die Wirtzelle einzuschleussen. Rechts: Botrytis EVs auf der Oberfläche einer Pflanzenzelle (elektronenmikroskopsiche Aufnahme erstellt zusammen mit Prof. A. Klingl, LMU München).
Aus dem Verständnis, wie Pathogene ihre RNAs in Wirtszellen einschleusen können, erwarten wir neue Möglichkeiten für die Entwicklung RNA-basierter Anwendungen im Pflanzenschutz (Cai et al., 2019).
Argonautes
Argonautes (AGO) sind RNA-bindende Proteine, die mittels RNA-Sequenzspezifität gezielt die Gen-Expression über RNAi unterdrücken können. AGOs sind strukturell und funktionell sehr konserviert und kommen sowohl in Pflanzen als auch in eukaryotischen Pathogenen vor. AGOs sind zentral für den Cross-Kingdom RNAi Mechanismus, da sowohl Pflanzen als auch Pathogene ihre RNAs in die AGOs des Interaktionspartners einschleusen (Cheng et al. 2025). Dabei interessieren wir uns für den Beladungsmechanismus von AGOs mit externen, nicht-eigenen RNAs.
Hierzu haben wir Methoden entwickelt, die Bindeeigenschaften von AGOs mit RNA zu untersuchen. Mittel Immuno-Capturing können wir z.B. gezielt pflanzliche AGOs während der Pathogen-Infektion aufreinigen, um neue Pathogen RNA-Effektoren mittels Hochdurchsatz-Sequenzierung zu identifizieren (Dunker et al. 2021).
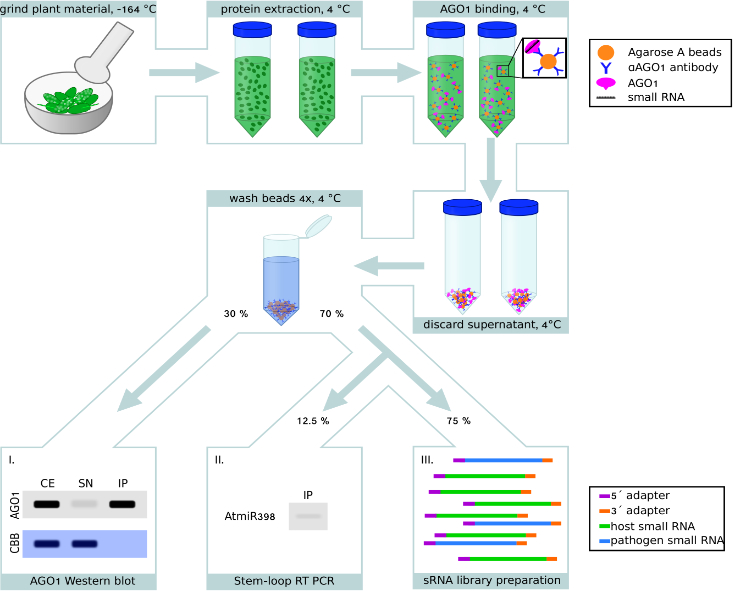
AGO Immuno-capturing aus infizierten Pflanzen
Retrotransposons und RNA Diversität
Der Pilz Botrytis cinerea produziert eine Vielzahl kleiner RNAs, die während der Infektion in die Pflanze eingeschleust werden, um die Immunantwort zu unterdrücken (Cross-Kingdom RNAi). Ein Großteil dieser kleinen RNAs wird von Retrotransposons kodiert. In einer Studie konnten wir zeigen, dass sich die Profile dieser Retrotransposon-RNAs in verschiedenen Botrytis-Isolaten stark unterscheiden (Porquier et al. 2021). Diese Unterschied korreliert mit der Virulenz des Pilzes. Wir sind daran interessiert zu erforschen, welchen Einfluss Retrotransposons und kleine RNAs auf die Virulenz und das Wirtsspektrum von pathogenen Pilzen hat.
![]()
Links: Botrytis-Isolate, mitte: kleine RNA-Sequenzierungsprofile, rechts: Virulenzunterschiede verschiedener Botrytis-Isolate auf Tomatenblätter.